Structure of bacteria
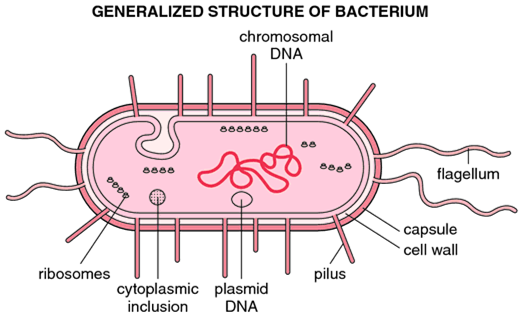
Cell wall
Prokaryotic cells almost always are bounded by a fairly rigid and chemically complex structure present between the cell membrane and capsule/slime layer called the cell wall. Peptidoglycan is the main component of the cell wall and is responsible for the shape and strength of the cell. It is a disaccharide and contains two sugar derivatives—N-acetylglucosamine and N-acetylmuramic acid—joined together by short peptide chains. N-acetylmuramic acid carries a tetrapeptide side chain consisting of D- and L-amino acids (D-glutamic acid and L-alanine) with mesodiaminopimelic acid (Gram-negative bacteria) or L-lysine (Gram-positive bacteria). Tetrapeptide side chains are interconnected by pentaglycine bridges. Most Gram-negative cell walls lack an interpeptide bridge. The cell wall provides shape to the cell and protects bacteria from changes in osmotic pressure, which within the bacteria cell measures 5–20 atmospheres.
Bacterial cells can be classified into Gram-positive or Gram-negative based on the structural differences between Gram-positive and Gram-negative cell walls. The cell walls of the Gram-positive bacteria have simpler chemical structures compared to Gram-negative bacteria.
Gram-positive cell wall
The Gram-positive cell wall is thick (15–80 nm) and more homogenous than that of the thin (2 nm) Gram-negative cell wall. The Gram-positive cell wall contains a large amount of peptidoglycan present in several layers that constitute about 40–80% of the dry weight of the cell wall. The Gram-positive cell wall consists primarily of teichoic and teichuronic acids. These two components account for up to 50% of the dry weight of the wall and 10% of the dry weight of the total cell.
1. Teichoic acids:
Teichoic acids are polymers of polyribitol phosphate or polyglycerol phosphate-containing ribitol and glycerol. These polymers may have sugar or amino acid substitutes, either as a side chain or within the chain of the polymer. Teichoic acids are of two types; wall teichoic acid (WTA) and lipoteichoic acid (LTA). They are connected to the peptidoglycan by a covalent bond with the six hydroxyls of N-acetylmuramic acid in the WTA and to plasma membrane lipids in LTA.
2. Teichuronic acid:
Teichuronic acid consists of repeat units of sugar acids (such as N– cetylmannuronic or D-glucuronic acid). They are synthesized in place of teichoic acids when phosphate supply to the cell is limited. The gram-positive cell wall also contains neutral sugars (such as mannose, arabinose, rhamnose, and glucosamine) and acidic sugars (such as glucuronic acid and mannuronic acid), which occur as subunits of polysaccharides in the cell wall.
Gram-negative cell wall
The Gram-negative cell wall is much more complex than the Gram-positive cell wall. Peptidoglycan content in the Gram-negative cell wall is significantly less than the Grampositive cell wall. Only 1–2 layers of peptidoglycan (2–8 nm) are present just outside the cell membrane. The Gram-negative cell wall outside the peptidoglycan layer contains three main components—(a) lipoprotein layer, (b) outer membrane, and (c) lipopolysaccharides.
Lipoprotein layer:
The lipoprotein layer is mainly composed of Braun’s lipoprotein. Braun’s lipoprotein is a small lipoprotein that is covalently joined to the underlying peptidoglycan and embedded in the outer membrane by its hydrophobic end. The lipoprotein stabilizes the outer membrane of the Gram-negative cell wall.
Outer membrane:
The outer membrane is a bilayered structure; its inner part resembles in composition with that of the cell membrane, while its outer part contains a distinctive component called lipopolysaccharide. The outer membrane and plasma membrane appear to be in direct contact at many sites in the Gram-negative wall. The outer membrane has a variety of proteins as follows:
(a) Porins:
The outer membrane has special channels consisting of protein molecules called porins. These porins have many functions:
■ They permit the passive diffusion of low-molecular-weight hydrophilic compounds, such as sugars, amino acids, and certain ions;
■ They exclude hydrophobic molecules; and
■ They serve to protect the cell.
(b) Outer membrane proteins (OMPs):
These include the following:
■ Omp C, D, F, and PhoE & LamB are the four major proteins of the outer membrane that are responsible for most of the transmembrane diffusion of maltose and maltodextrins.
■ Tsx, the receptor for T6 bacteriophage, is responsible for the transmembrane diffusion of nucleosides and some amino acids.
■ Omp A protein anchors the outer membrane to the peptidoglycan layer. It is also the sex pilus receptor in F-mediated bacterial conjugation. The outer membrane also contains proteins that are involved in the transport of specific molecules, such as vitamin B12 and iron-siderophore complexes; it also contains a limited number of minor proteins, such as enzymes, phospholipases, and proteases.
Lipopolysaccharides:
Lipopolysaccharides (LPS) are complex molecules present in the outer membrane of Gram-negative bacteria. Structurally, the LPS consists of three main components; lipid A, the core oligosaccharide, and the O polysaccharide or O-antigen.
■ Lipid A:
This consists of phosphorylated glucosamine disaccharide units, to which a number of long-chain fatty acids are attached. This also consists of hydroxymyristic acid, a unique fatty acid, which is associated with the endotoxic activity of the LPS. There is a little variation in the structure of the lipid A among different species of Gram-negative bacteria. However, it remains the same within the bacteria of the same species.
■ Core oligosaccharide:
The core oligosaccharide includes two characteristic sugars; keto-deoxy octanoic acid (KDO) and a heptose—both joined together by lipid A. This is genus-specific and similar in all Gram-negative bacteria. Lipooligosaccharides (LOS) are smaller glycolipids. They have relatively short, multiantennary (i.e., branched) glycans present in bacteria (e.g., Neisseria meningitidis, N. gonorrhea, Haemophilus influenzae, and Haemophilus ducreyi) that colonize mucosal surfaces. They exhibit extensive antigenic and structural diversity even within a single strain. LOS is an important virulence factor. Epitopes on LOS have a terminal N-acetyllactosamine residue, which is immunochemically similar to the precursor of the human erythrocyte i antigen. Sialylation of the N-acetyllactosamine residue in vivo provides the organism with the environmental advantages of molecular mimicry of a host antigen and the biologic masking thought to be provided by sialic acids.
■ O polysaccharide or O-antigen:
It is the portion extending outwards from the core. It has several peculiar sugars and varies in composition between bacterial strains, conferring species-specific antigen specificity. It is exposed to the host– immune system. Gram-negative bacteria may thwart host defenses by rapidly changing the nature of their O side chains to avoid detection.
Periplasmic space
Periplasmic space is a distinct space between the cell membrane and outer membrane (innermost layer of Gram-negative cell wall) in Gram-negative bacteria. This space is filled with a loose layer of peptidoglycan matrix. The periplasmic space of Gram-negative bacteria contains many proteins that participate in nutrient acquisition, and many hydrolytic enzymes, beta-lactamases binding proteins, and enzymes that participate in the peptidoglycan synthesis. Polymers of D-glucose, called membrane-derived oligosaccharides, appear to play a role in osmoregulation. The periplasmic space is less distinct in Gram-positive cell walls.

Cell wall of acid-fast bacilli
The cell wall of acid-fast bacilli, such as M. tuberculosis, contains large amounts of waxes known as mycolic acids. The cell wall is composed of peptidoglycan and an outer asymmetric lipid bilayer. The inner lipid bilayer contains mycolic acids linked to an arabinoglycan protein and the outer layer contains other extractable lipids. This hydrophobic structure renders these bacteria resistant to many harsh chemicals including detergents and strong acids. During staining, if the dye is introduced into these cells by brief heating or treatment with detergents, they resist decolorization by sulfuric acid or acid alcohol and are therefore called acid-fast organisms.
Cell membrane
Cell membrane or plasma membrane is a thin (5–10 nm) semipermeable membrane that acts as an osmotic barrier. It lies beneath the cell wall separating it from the cell cytoplasm. Cell membrane primarily contains phospholipids and proteins. It also contains enzymes associated with DNA biosynthesis, cell wall polymers, and membrane lipids. Bacterial plasma membranes usually have a higher proportion of protein than eukaryotic membranes. They usually differ from eukaryotic membranes in lacking sterols, such as cholesterol, except in Mycoplasma. The cell membrane has the following functions:
■ It acts as a semipermeable membrane regulating the inflow and outflow of metabolites to and from the protoplasm.
■ It helps in electron transport and oxidative phosphorylation.
Cytoplasm
Bacterial cytoplasm is a colloidal suspension of a variety of organic and inorganic solutes in a viscous watery solution. The matrix is largely formed by nearly 70% water. The cytoplasm contains all the biosynthetic components required by a bacterium for growth and cell division, together with genetic material. Prokaryotic cytoplasm, unlike that of eukaryotes, lacks endoplasmic reticulum and mitochondria. It also does not show any protoplasmic streaming. Bacteria lack a true cytoskeleton. The cytoplasm consists of ribosomes, mesosomes, and intracytoplasmic inclusions bodies.
Ribosomes:
The cytoplasmic matrix often is packed with ribosomes. Ribosomes look like small, featureless particles at low magnification in electron micrographs. They are smaller than their eukaryotic counterpart with the sedimentation of the 70S, compared with 80S in eukaryotes. They consist of two subunits of the 30S and 50S, giving a net 70S. Ribosomes are important because:
■ They serve as the sites of protein synthesis; matrix ribosomes synthesize proteins destined to remain within the cell, whereas plasma membrane ribosomes make proteins for transport to the outside.
■ They are also the sites of actions of several antibiotics, such as aminoglycosides, macrolides, and tetracyclines.
Mesosomes:
These are vesicular convoluted or multi-laminated structures formed as an invagination of the plasma membrane into the cytoplasm. Mesosomes are of two types—septal and lateral. The septal mesosome attached to the bacterial DNA is believed to coordinate nuclear and cytoplasmic divisions during binary fission. The function of lateral mesosomes still remains to be known. Mesosomes are analogous to the mitochondria of eukaryotes and are the principal sites of respiratory enzymes in bacteria.
Intracytoplasmic inclusion bodies:
Intracytoplasmic inclusion bodies are present in the protoplasm of bacteria. Their main function is believed to be storage. This occurs when their main constituent element is present in excess in the culture medium. Since inclusion bodies are used for storage, their quantity can vary depending on the nutritional status of the cell. They are the sources of carbon, inorganic substances, and energy. Some inclusion bodies also function to reduce osmotic pressure.
They may be of two types:
(i) organic inclusion bodies, which usually contain either glycogen or polyhydroxy butyrate, and
(ii) inorganic inclusion bodies, which may be of polyphosphate granules or sulfur granules.
Examples of intracytoplasmic inclusion bodies include metachromatic granules or volutin granules, starch inclusions, and lipid inclusions. Volutin granules, typically present in C. diphtheriae, can be demonstrated by Albert’s stain. Similarly, starch granules present in the bacteria can be demonstrated on staining with iodine. Lipid inclusion found in Mycobacteria is demonstrated by Sudan black dye.
Nucleus
The bacterial nucleus is neither enclosed in a nuclear membrane nor associated with any nucleolus. It is a haploid and replicates by simple fission. The nucleus of the bacteria consists of a single circle of double-stranded deoxyribonucleic acid (DNA), arranged in a supercoiled circular structure. It measures about 1000 μm when straightened. The chromosome is located in an irregularly shaped region called nucleoid but is often referred to as a bacterial chromosome because of the analogy with the eukaryotic structure. The nucleoid is visible through the light microscope after staining with the Feulgen stain, which specifically reacts with DNA. In actively growing bacteria, the bacterial DNA can account for up to 20% of the volume of the bacterium and has projections that extend into the cytoplasmic matrix. Careful electron microscopic studies often have shown the nucleoid to be in contact with either the mesosome or the plasma membrane.
Many bacteria also possess smaller circles of extrachromosomal DNA called plasmids. The plasmids are double-stranded DNA molecules, usually circular, that can exist and replicate independently. Plasmids are not required for host growth and reproduction, although they may carry genes that confer the bacterium with properties such as antibiotics resistance or the capacity to produce toxins or enzymes.
Capsule
The capsule is mostly made up of polysaccharides, often referred to collectively as exopolysaccharides. Exopolysaccharides are sometimes neutral homopolysaccharides (e.g., the glucans and fructans of many oral streptococci) or negatively charged. However, Bacillus anthracis has a capsule comprising of polyamino acids, such as D-glutamic acid. The D-glutamic acid is probably analogous to the negatively charged polysaccharide capsule.
Slime layer
The Slime layer (S-layer) is a structured paracrystalline protein layer shown by electron microscopy. These are generally composed of a single kind of protein molecule, sometimes with carbohydrates attached. They are resistant to proteolytic enzymes and protein-denaturing agents. The slime layer protein protects the cell from wall-degrading enzymes and bacteriophages. It plays an important role in the maintenance of cell shape, and it may be involved in cell adhesion to host epidermal surfaces.
Flagella
Bacterial flagella are thread-like appendages intricately embedded in the cell envelope. These structures are responsible for conferring motility to the bacteria. The arrangement of flagella varies between different bacterial species. Depending on the arrangement, flagella can be of the following types:
■ Monotrichous (single polar fl agellum), e.g., Vibrio cholerae.
■ Lophotrichous (multiple polar fl agella), e.g., Spirilla.
■ Peritrichous (flagella distributed over the entire cell), e.g., Salmonella Typhi, E. coli, etc.
■ Amphitrichous (single flagellum at both the ends), e.g., Spirillum minus.

Structure:
The flagella are 3–20 μm in length and 0.01–0.03 μm in diameter. The main part of the filament is made up of protein subunits called flagellin arranged in several helices around a central hollow core. The flagellum is attached to the bacterial cell body by a complex structure consisting of a hook and a basal body. The basal body bears a set of rings, one pair in Grampositive bacteria and two pairs in Gram-negative bacteria, through which the bacteria rotate either in a clockwise or an anticlockwise direction. Above the base of the filament is the hook, a short curved structure between the external filament and basal body. This part produces a propeller-like repulsion from the revolving flagellum. Spirochetes are motile bacteria but without any external flagella. They are motile due to the presence of an axial filament. Axial filament consists of a bundle of flagellum-like structures that lie between the cell surface and an outer sheath and connect one end of the cell to the other. They are sometimes called the endoflagellates.
Function:
Flagella have the following functions:
■ They are primarily responsible for the motility of bacteria by chemotaxis.
■ They may play a role in bacterial survival and pathogenesis. They are highly antigenic, they possess H antigens, and some of the immune responses to infection are directed against these proteins. The flagella of different bacteria differ antigenically. Flagellar antibodies are not protective but help in serodiagnosis.
Pili (fimbriae)
Pili or fimbriae are synonymous for most purposes. They are hair-like filaments that extend from the cell surface and are found almost exclusively on Gram-negative bacteria. They are composed of structural protein subunits termed pilins. Minor proteins termed adhesins are located at the tips of pili and are responsible for the attachment properties.
Structure:
The pili are shorter and straighter than flagella, although the basic structure is the same. Like flagella, it consists of helices of a protein called pilins, arranged around a hollow core but without a motor. They are 0.5 μm long and 10 nm thick. They are antigenic in nature. Pili hemagglutinate RBCs of guinea pigs and are specifically inhibited by mannose, on the basis of which they are classified into four types as follows:
- Type 1:These occur in E. coli, Klebsiella, Shigella, and S almonella. They are mannose sensitive.
- Type 2:These are present in Salmonella Gallinarum and Salmonella Pullorum, devoid of any hemagglutinating or adhesive properties.
- Type 3:These are present in some strains of Klebsiella, S erratia, etc. They agglutinate RBC only after heating and are mannose resistant.
- Type 4: These are mannose resistant and occur in Proteus.
Sex pili:
A specialized kind of pili called sex pili is responsible for the attachment of donor and recipient cells in bacterial conjugation. These pili are longer (10–20 μm) and vary 1–4 in number. The sex pili are of two types:
(i) F pili: They specifically adsorb male-specific RNA and DNA bacteriophages. They are encoded by sex factor F and fertility inhibition–positive resistance factors (fi + R factors).
(ii) I pili: They adsorb male-specific filamentous DNA phages, encoded by col factor and fi -R factor.
Function:
- Adherence of symbiotic and pathogenic bacteria to host cells, which is a necessary step in initiation of infection.
- Transfer of bacterial DNA takes place through sex pili during the process of conjugation.